Article Content
Introduction
Nanopore sensing based on transmembrane proteins has exhibited great potential in the field of label-free and highly sensitive single-molecule detection1,2,3,4,5. Conventional protein nanopores such as α-HL, MspA, and aerolysin have made great progress in DNA and protein sequencing6,7,8, amino acid recognition9,10, biomarker detection11,12,13,14,15, and single-molecule interactions study16,17,18,19. The development of new biological pores is considered a crucial means to enhance the resolution and expand the detection range of analytes. The membrane-insertion capacity (surface hydrophobicity) and ionic transport property (inside hydrophilicity) are two important indicators to evaluate whether a channel structure is suitable for constructing a nanopore sensor. Under this principle, some novel barrel-shaped nanopores including α-helical peptide pores20,21,22, β-barrel nanopores23,24, and DNA-based nanopores25,26,27 were designed and constructed. Usually, to enhance the hydrophobicity, the de novo pore design necessitates complex chemistry involving unnatural residues or tags incorporation. In a traditional perspective, the barrel structured channel with hydrophobic surface and hydrophilic inwall28,29 is preferred in the development of novel nanopores. However, natural proteins with non-barrel shape but possess promising membrane-insertion capacity and ionic transport properties remain largely unexplored.
Ferritin is a globular protein composed of 24 subunits with the function of storing and releasing iron to maintain the iron balance in living organisms30,31,32,33. To our best knowledge, the ferritin, with large size and globular configuration, was never reported to be inserted into the lipid bilayer and function as a transmembrane channel, let alone for single-molecule sensing. However, viewing from membrane-insertion capacity, the hydrophobic sites including six four-fold (C4) channels on the surface of ferritin could provide the attachment points for phospholipid molecules. On the other hand, ferritin has eight hydrophilic three-fold (C3) channels with narrow aperture (3-5 Å), which are considered as the natural ion channels for iron transport34,35, also providing the confined space for potential single-molecule sensing. Besides, there are three natural cysteine residues inside C3 channel34, which provide the possible reactive sites for single-molecule interaction, avoiding the complicated protein mutation during nanopore engineering. Thereby, we consider ferritin as an excellent candidate for a transmembrane protein to build a novel high-resolution nanopore sensor.
Here, we successfully insert single horse spleen ferritin into the planar lipid bilayer, constructing the ferritin-based transmembrane nanopore. The multiple insertion experiments and molecular dynamics (MD) simulation are used to confirm the establishment of a single-ferritin transmembrane system and evaluate the stability of the ferritin-lipid bilayer system, respectively, which indicates the ferritin is able to be individually and stably embedded in the lipid bilayer. The asymmetric I-V curves and voltage-dependent channel opening and closing behavior are discovered, revealing the ionic current rectification and voltage-gating characteristics of the ferritin nanopore. Furthermore, the ferritin nanopore is developed as a single-molecule sensor for amino acid and peptide detection with the assistance of Cu2+. The detection mechanism is also explored by combining controlled nanopore experiments and multiscale MD simulations. The Cu2+ is demonstrated to bridge analytes and residues (Cys126 and Glu130) inside the ferritin C3 channel, leading to the blockage current signals. The globular ferritin nanopore confirms that a high-resolution nanopore sensor could be constructed by non-barrel protein which greatly expands the protein candidates as nanopore sensors.
Results and discussion
Electrical properties and MD simulation of ferritin nanopore on the lipid bilayer
The horse spleen ferritin is assembled by 24 identical subunits, resulting in a polyhedron-like globular structure with a size of ~10 nm, as demonstrated in the previous works32,36,37. There are eight hydrophilic C3 channels on the vertex of ferritin and six hydrophobic C4 channels on the face center of ferritin (Fig. 1a, b). Specifically, the C3 channel is comprised of three subunits, in which three-fold Cys126, Asp127, and Glu130 residues enable a pore with the size of 3-5 Å (Fig. 1a). The C3 channel is considered as the specific channel through which ferritin absorbs and releases Fe2+. In detail, the Cys126 residue near the external entrance of C3 channel is reported to play a key role in absorbing Fe2+ 34. The C3 ionic channel makes it possible to construct a ferritin nanopore with stable ionic current. To be noted that the ferritin sample used in nanopore experiments (Sigma-Aldrich) is comprehensively characterized by SEC, SDS-PAGE, Native-PAGE, DLS, and HRTEM experiments and identified as intact L-rich ferritin with spherical structure (Supplementary Fig. 1). For convenience, the “ferritin” was used to represent the L-rich horse spleen ferritin in this work.
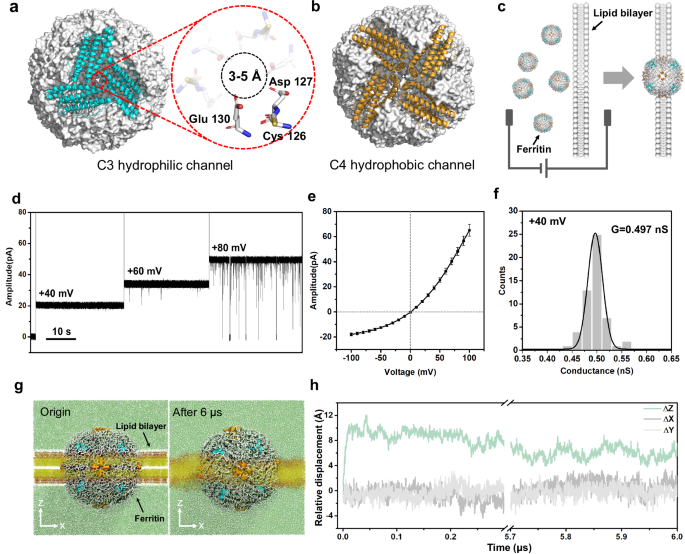
a, b Hydrophilic C3 channel (blue) and hydrophobic C4 channel (orange) of ferritin (grey). The C3 channel with the size of 3–5 Å is surrounded by three-folds of Cys126, Asp127, and Glu130, which plays the key role in ion transport. c Cartoon models of inserting ferritin nanopore into the lipid bilayer. d Electrical recording of ferritin nanopore showing voltage-dependent channel opening and closing. e I − V curve obtained from a single ferritin nanopore with nine independent experiments. The centre and error bar represent mean value and standard deviation (SD), respectively. f Histogram and Gaussian fitting of channel conductance obtained from 53 independent single-ferritin recordings at +40 mV. g The multiscale model of ferritin-lipid bilayer system before (left) and after (right) MD equilibrium. The fatty tails and the head group of 1,2-diphytanoyl-sn-glycero-3-phosphocholine (DPhPC) are colored yellow and red, respectively. Water molecules are marked green. h Relative displacement along three directions between the mass center of ferritin and all phospholipid molecules.
The ferritin nanopore sensing apparatus is prepared by inserting one ferritin molecule into the planar lipid bilayer, where four C4 channels are embedded in the lipid bilayer, and eight C3 channels are distributed on both sides of the membrane, with four on each side. (Fig. 1c and Supplementary Fig. 2). The abrupt current increasing shown in Supplementary Fig. 3 indicates the successful construction of ferritin nanopore. The consistent pore-forming currents in three independent nanopore insertion experiments demonstrate the reliability of the ferritin nanopore (Supplementary Fig. 4). After that, the electrical properties of the ferritin nanopore are characterized. The voltage-gated characteristic of ferritin nanopore is discovered, which maintains the open state at a low voltage of +40 mV but starts to randomly close at a high voltage of +80 mV (Fig. 1d). To be noted that, all the electrophysiological recordings in this work, if without special illustration, are carried out at +40 mV to avoid the random opening-closing background signals, despite the background signals at higher voltage of +80 mV have insignificant interference in detecting analytes (Supplementary Fig. 16). In addition, the asymmetric I-V curve of ferritin nanopore is observed, exhibiting larger conductance at positive voltages and smaller conductance at negative voltages (Fig. 1e). Several insertion experiments exhibit the same I-V curves with a specific current of ~65 pA at +100 mV and ~-18 pA at -100 mV, revealing the uniformity of ferritin nanopore. (Fig. 1e and Supplementary Fig. 5). The further insertion of a second ferritin causes a double ionic current and keeps the specific I-V characteristic of ferritin nanopore (Supplementary Fig. 6), revealing that the special I-V curve shown as Fig. 1e means only one ferritin was inserted into the membrane. The ferritin nanopore has a mean conductance (G) of 0.497 ± 0.045 nS at +40 mV, based on the statistical analysis of n = 53 independent experiments (Fig. 1f). This conductance corresponds to ~1.25 × 108 ions per second, which is consistent with the magnitude of ion channels38. The ionic current is believed to result from ions passing through only C3 channels rather than hydrophobic C4 channels, according to the reported ions transport pathway of ferritin34. The above asymmetric I-V curve, voltage-gating behavior, and high ion flux are the features of ion channels38,39,40, confirming the successful construction of a single ferritin nanopore on the lipid bilayer. It is worth noting that the L-subunit of ferritin appears to play a key role in facilitating the embedding of ferritin into lipid bilayer. In our experiments, both L-rich horse ferritin and recombinantly expressed human L-ferritin (LFn) could successfully embed into lipid bilayer, whereas recombinantly expressed human H-ferritin (HFn) consistently failed to incorporate despite multiple attempts.
To evaluate the stability of the ferritin-lipid bilayer system, the coarse-grained molecular dynamics (CG-MD) simulation41,42,43,44,45,46 is employed. (Please refer to Methods for modeling details). The initial conformation is shown on the left side of Fig. 1g. The ferritin is placed at the center of the 1,2-diphytanoyl-sn-glycero-3-phosphocholine (DPhPC) bilayer, and the four C4 hydrophobic channels are located on the XY plane in the middle of the lipid bilayer. The system is then relaxed by the isobaric-isothermal ensemble (NPT ensemble)47,48 for 6.0 μs, with the reference temperature set to 373 K to speed up conformational changes. For the first 0.05 μs, the embedding ferritin exhibits obvious movement relative to the lipid bilayer on the Z-direction. As shown in Fig. 1h, the mass center of ferritin moves away from that of the bilayer along the Z-direction and quickly reaches equilibrium, meanwhile having almost no displacement along the X and Y axes, indicating that the ferritin can be stably embedded in the lipid bilayer. The final conformation is shown on the right side of Fig. 1g. In the simulation system, four of six hydrophobic C4 channels are always embedded in the phospholipid membrane and all eight hydrophilic C3 channels are exposed on both sides of the bilayer, directly contacting with the solution. The C3 channels exposed in the solution are considered to be the channels leading the ionic current.
Single-molecule sensing with ferritin nanopore
The dimensions of most common protein nanopores are larger than 7 Å49, while the C3 channel of ferritin has a smaller channel size (3-5 Å), which could render a higher spatial resolution. In addition, the cysteine mutation is most used in nanopore engineering to enhance the reactivity of nanopores50,51,52, while there are three native cysteines in the C3 channel, enabling ferritin nanopore a good reactivity even without mutation. Therefore, the ferritin nanopore can be developed as a single-molecule sensor with high resolution and reactivity. Both L-Cysteine (L-Cys) and L-Homocysteine (L-Hcy) are significant biothiols that are considered as biomarkers related to colorectal cancer and cardiovascular diseases53,54. However, the slight difference (only one methylene) between L-Cys and L-Hcy causes similar physical and chemical properties, leading to a difficult discrimination between them. Lu et al. successfully differentiated L-Cys and L-Hcy assisted by synthetic complex probe with mutant AeL nanopore55. Cao et al. also achieved the differentiation between them by introducing [AuCl4]– as a bridge with methionine-mutated MspA nanopore, in which the sulfur atom on methionine plays a key role56. With three reactive cysteine residues in each C3 channel, the ferritin nanopore is considered as a proper sensor for the discrimination between L-Cys and L-Hcy.
Based on as-constructed ferritin nanopore sensor, direct measurement of L-Cys (80 μM) in the KCl buffer (1 M KCl, 10 mM MOPS, pH 7.4) under +40 mV bias results in no obvious blockage events (Supplementary Fig. 7). Then, the Cu2+ is added as connector because of its capability to form Cys-Cu five-membered ring complex and bind with the Cys126 residue on the C3 channel34,57. So, we design to sense L-Cys and L-Hcy with the assistance of Cu2+. The mixture solutions of L-Cys (80 μM) and L-Hcy (80 μM) are respectively prepared in the KCl buffer with Cu2+ (80 μM). Interestingly, the homogeneous current blockage signals are observed in both L-Cys and L-Hcy mixture solutions at +40 mV (Fig. 2b, c). In detail, the L-Cys leads to smaller current blockage signals (ΔI/I0 = 0.331) with longer dwell time and the L-Hcy leads to larger current blockage signals (ΔI/I0 = 0.388) with shorter dwell time (Fig. 2d, e), in which the disparity in ΔI/I0 for them is attributed to the different sizes of thiol-Cu complexes. The Cys and Hcy events are simultaneously observed in the continuous recording, displaying the distinct discrimination (Supplementary Fig. 8). It is clear that the good resolution between L-Cys and L-Hcy is achieved by our non-engineered ferritin nanopore, although there is only a slight difference by one methylene between them. In addition, multi-level blockage signals are observed both in the detection of L-Cys and L-Hcy, which is considered as the simultaneous sensing for multiple molecules by the multi-channel ferritin sensor (Supplementary Fig. 9a). The multi-level signal of L-Cys, as an example, is analyzed, in which the quantitative relationship between the probability of multi-level signals and the Cys-Cu concentrations is described using binomial distribution model and logistic model (Supplementary Fig. 9b–d). At the same time, compared with ferritin, the apo-ferritin without internal iron core exhibits the identical sensing behavior for Cys-Cu detection, revealing that the conductive and sensing properties of ferritin nanopore are independent of the iron core loaded in ferritin, but attributed to the multi-channel structure and the suitable sensitive sites in the channels (Supplementary Fig. 10).
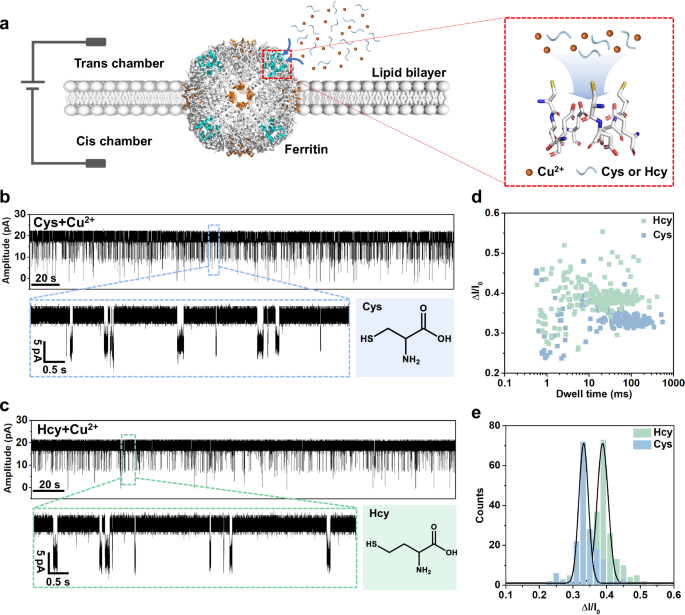
a Schematic of ferritin nanopore sensor. The lipid bilayer divides the flow cell into cis (grounded) and trans chambers, and the testing solution is added into the trans chamber for detection. b Current trace at +40 mV in the presence of L-Cys (80 µM) and Cu2+ (80 µM) in the trans side. c Current trace at +40 mV in the presence of L-Hcy (80 µM) and Cu2+ (80 µM) in the trans side. d, e Scatter plot and histogram showing the dwell time and ΔI/I0 of blockage signals in Cys (n = 174) and Hcy (n = 201) sensing.
In addition, to test the qualification capacity of ferritin nanopore, the qualification experiments for L-Cys are carried out. The L-Cys concentration standard curve is fitted according to the acquired event frequency from standard solutions. (Supplementary Fig. 11a) Then, the actual sample (L-cysteine capsule) is dissolved and detected in three independent nanopore experiments. According to the fitted concentration standard curve, we acquired the calculated L-cysteine weight percentage as 16.6 wt%±1.9 wt%, close to the indicated value (18 wt%), revealing the well qualification capacity of the ferritin nanopore (Supplementary Fig. 11b).
Mechanism exploration for cysteine sensing with ferritin nanopore
The key role of Cu2+ in the formation of Cys-Cu complex is explored, in which the factors of chelator, buffer, and pH are investigated. The pure Cu2+ (80 μM) is firstly tested and there is no obvious signal appearing, indicating the Cu2+ itself does not induce blockage signals (Fig. 3a). Then, the key role of Cu2+ in the sensing process is further verified by the in-situ nanopore controlled experiments. There are no homogeneous signals appearing with only L-Cys, however, after adding Cu2+, the homogeneous blockage events are observed (Fig. 3b). To be noted that, the blockage signals were gradually observed after ~160 s rather than immediate appearance. We guess that in this period, the L-Cys combines with Cu2+ forming Cys-Cu complex and causing the characteristic signals. Furthermore, after adding the excess Na4EDTA (160 μM) into the mixture to capture Cu2+, the blockage events disappear, revealing the Cu2+ is a key factor in the sensing process (Fig. 3c). The influence of buffer reagents and pH of the testing solution is also evaluated. It is clear that the events frequency decreases after only replacing the MOPS with Tris-HCl (Fig. 3d, and Supplementary Fig. 12), which is attributed to the strong binding between metal ions and Tris-HCl58, interfering with the formation of Cys-Cu complex. In addition, the dwell times of blockage signals shorten as pH decreases, which is consistent with the weaker interactions between Cu2+ and L-Cys at lower pH (Fig. 3e and Supplementary Fig. 13)59. Besides, other metal ions including Fe3+, Fe2+, Zn2+, and Ni2+ were also explored as connectors for sensing L-Cys, however, no blockage signals were observed, implying the specificity of Cu2+ in this system (Supplementary Fig. 14). All the abovementioned experiments reveal that the coordination between Cu2+ and Cys may play the key role in causing blockage events.
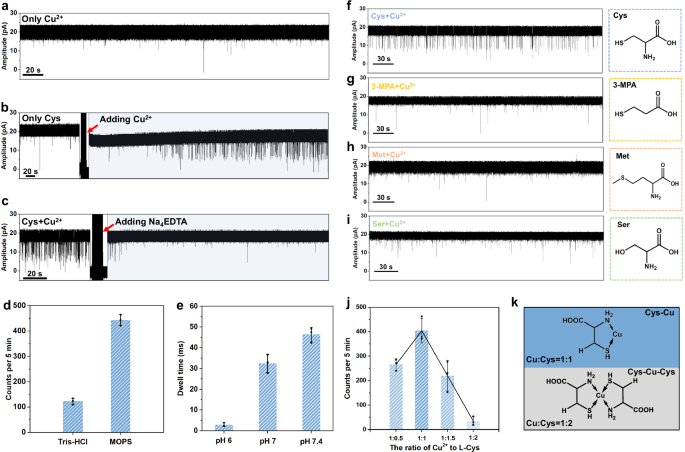
a Only Cu2+ (80 µM) shows no obvious blockage signals. b Current trace of Cys (80 µM) before and after the in-situ addition of Cu2+ (80 µM). c Current trace of Cys (80 µM) and Cu2+ (80 µM) before and after the in-situ addition of Na4EDTA (160 µM). d Effects of buffers on event frequency of Cys (80 µM) and Cu2+ (80 µM). The error bar comes from three different time segments. e Influence of pH on the dwell time of Cys (80 µM) and Cu2+ (80 µM). The error bar comes from three independent experiments. f–i The controlled experiments for cysteine analogues with different functional groups, showing the essential role of -SH and -NH2 on cysteine to generate blockage events. j The events frequency with the different binding ratios of Cu2+ to Cys. The error bar comes from three independent experiments. k Proposed structures of Cys-Cu and Cys-Cu-Cys complexes with the different binding ratios of Cu2+ to Cys (1:1 and 1:2). For all the bar charts in this figure, the centre and error bar represent mean value and standard deviation (SD), respectively.
To further explore the binding state of Cys-Cu complex, the quantum chemical calculation is used to screen the possible structures, including N-Cu-O, S-Cu-O, and S-Cu-N. The B3LYP-D3/6-31 g(d) method60,61 and XYGJOS/def2-TZVPP method62,63,64 are used to optimize conformations and calculate the relative internal energies of three structures, respectively. As a result, the S-Cu-N five-membered ring binding state is considered as the most stable state for Cys-Cu complex due to its lowest relative internal energy (Supplementary Table 2). In addition, the nanopore controlled experiments are also performed for cysteine analogues, including 3-Mercaptopropionic acid (3-MPA), L-Methionine (L-Met), and L-Serine (L-Ser), which contain only one different functional group comparing to L-Cys or L-Hcy (Fig. 3f-i). As a result, no obvious blockage signals under the same testing conditions were observed for three counterparts, confirming that the synergistic binding of amino (-NH2) and sulfhydryl (-SH) with Cu2+ plays a key role in the formation of S-Cu-N structure to generate characteristic blockage signals. It is worth noting that Cu²⁺ has previously been reported as connectors to help detect amino acids in nanopore methods, where amino acids are thought to bind to Cu2+ via their amino and carboxyl groups9,65. For example, Zhang et al. utilized MspA nanopore to detect 20 amino acids with the aid of Cu2+ 9. However, the ferritin C3 channel is smaller than the MspA nanopore, leading to a stronger electric field at the entrance. Thus, only the form of most stable S-Cu-N rather than N-Cu-O strongly remains in the channel and generates signals. The sulfhydryl group on analytes is necessary for nanopore sensing with the ferritin nanopore. It is reported that the Cu2+ is able to bind one or two cysteine molecules to form different Cys-Cu complexes (Cys-Cu and Cys-Cu-Cys)57,66. To explore the influence of the binding ratio of Cu2+ to Cys, the solutions with different ratios of Cu2+ to Cys (1:0.5, 1:1, 1:1.5, and 1:2) are prepared and tested (Fig. 3j and Supplementary Fig. 15). To be noted that the concentration of Cu2+ is fixed at 80 µM, while the concentrations of Cys are set at 40, 80, 120, and 160 µM, corresponding to the ratios of 1:0.5, 1:1, 1:1.5, and 1:2, respectively. When the ratio of Cu2+ to Cys is 1:0.5, the events frequency is calculated to ~265 events per 5 min. After increasing the concentration of L-Cys to the ratio of 1:1, the events frequency improves to ~404 per 5 min. However, with the continued increase of L-Cys, the events frequency starts to decrease and the events are hardly observed with a frequency of ~35 per 5 min in the ratio of 1:2 for Cu2+ to L-Cys. According to the experimental results, we speculate that the Cu2+ binding with only one cysteine has remaining coordination sites to bind with the residues in the ferritin C3 channel causing the blockage signals, however, the Cu2+ chelated by two cysteines has no other coordination sites to interacts with residues in C3 channel resulting in no signals generation (Fig. 3k).
To further explore how the Cys-Cu complex binds with the ferritin C3 channel, a long-time (1 ns) quantum mechanics (QM)/all-atom (AA)/ coarse-grained (CG) MD simulation is carried out. The multiscale model is first constructed. The Cu2+, its coordinated residues, and active water molecules are represented by the level of QM with the GFN2-xTB semiempirical method67,68. Residues and water molecules near the QM region are defined as the AA region. Other residues and solvents far from the QM region are defined as CG regions. Figure 4a and 4b show the schematic and structural representations of the multiscale model. (Please refer to Methods for more modeling details). To evaluate the reasonability of our multiscale model, the interaction between single Cu2+ and residues in the C3 channel is simulated and compared to that in the crystal structure 3RE734,69. The observed coordination pattern of Cu2+ with His114, Cys126, and Glu130 in the simulation is similar to that in the crystal structure, confirming the reasonability of our multiscale model (Supplementary Fig. 17).
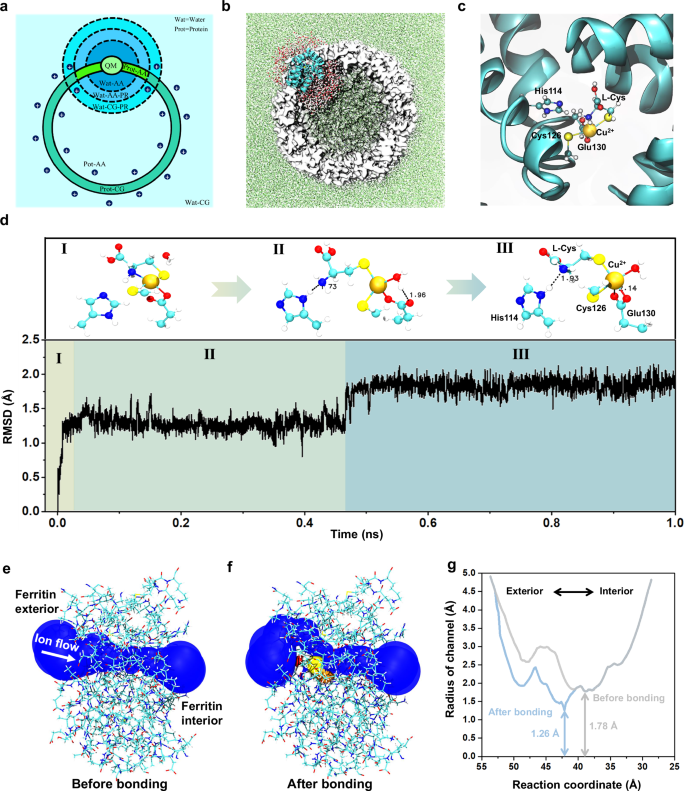
a A schematic diagram of the CG division map for different regions. Cys-Cu-Ferritin (Prot) is represented by three scales, QM, AA, and CG. Water solvents (Wat) are represented by two scales, AA and CG. Position restrictions (PR) are adopted for molecules near interfaces to prevent diffusion. b The initial conformation of the active and non-active region in the Cys-Cu-Ferritin. The active region corresponds to the AA & QM regions in a represented by the cyan cartoon (protein) and AA water molecules. Non-active CG regions are depicted using white surfaces (protein) and green dots (solvents), respectively. c The initial conformation of the QM region represented by Cu2+, L-Cys, His114, Cys126, and Glu130 and two nearby water molecules. d The root mean square deviation (RMSD) of Cu2+ and five coordinated atoms from residues with respect to the simulation time. Primary coordination patterns of corresponding RMSD are given above, with hydrogen bonds represented by doted lines. e, f The side view showing the shape of the ion transferring pore before and after Cys-Cu attached to the C3 channel. g The estimated radius of the channel before and after bonding Cys-Cu complex with respect to the reaction coordinate of ion flow. The reaction coordinate indicates the position of the ion flow within the C3 channel, with larger values corresponding to locations nearer to the exterior of the ferritin.
Then, the binding state of Cys-Cu complex in C3 channel is explored with QM/AA/CG multiscale MD simulation. First, the Cys-Cu complex is docked into the protein channel in a manner similar to the Cu(II) coordination in the crystal structure 3RE7. Cu(II) adopts a square pyramidal coordination mode, with two sulfur atoms in a trans position to reduce steric hindrance, the coordination pattern is shown in Fig. 4c. Afterwards, the Cys-Cu-Ferritin complex is solvated and coarse-grained to the QM/AA/CG multiscale system. The MD simulation is carried out for 1 ns to evaluate the stability of the coordination. The coordination pattern of the Cys-Cu-ferritin complex undergoes two major transitions (step I to II and step II to III) during the simulation, as shown in Fig. 4d and Supplementary Fig. 18. The coordination between -SH on Cys126 and Cu(II) always remains stable and the two oxygen atoms from -COOH on Glu130 alternately bind to the Cu(II), indicating both the Cys126 and Glu130 on the channel have stable interaction with Cu(II). As for the L-Cys, its -SH keeps stable interaction with Cu(II) but the -NH2 gradually moves away from Cu(II) and forms a stable hydrogen bond with the imidazole group on His114. In this way, the Cys-Cu complex is captured and stabilized in the C3 channel with multiple interactions from residues of His114, Cys126, and Glu130. To further verify the key roles of these residues, we introduce excess Zn2+, that also has stronger interaction with these residues34, in the detection for Cys-Cu complex to occupy these binding sites and analyze how the Cys-Cu events frequency becomes before and after introducing Zn2+. The MD simulation was first conducted to further confirm the coordination style of Zn2+ in C3 channel, in which, the Zn2+ was observed to indeed bind with these three residues and keep stable (Supplementary Fig. 19a, b). Furthermore, the concentration control experiments for Zn2+ were conducted, and it was found that the frequency of Cys-Cu events gradually decreased as the Zn2+ concentration increased, accordingly revealing the key roles of these residues (His114, Cys126, and Glu130) in binding with Cys-Cu complex (Supplementary Fig. 19c). In addition, the C3 channel radiuses before and after bonding Cys-Cu complex are calculated by MolAICal software70 (Fig. 4e–g). After bonding Cys-Cu complex, the channel radius at the narrowest point shows an obvious reduction, which causes the smaller ionic current, indicating the Cys-Cu complex is the key analyte in causing current blockage signals. Combining with the above controlled nanopore experiments and MD simulations, we result that the -NH2 and -SH of L-Cys play a key synergistic effect in chelating Cu2+ to form Cys-Cu complex. The Cys-Cu complex will enter into the ferritin C3 channel and then bind with His114, Cys126, and Glu130, causing the blockage events.
Dipeptides sensing with ferritin nanopore
To further explore the sensing applications, we attempt to detect cysteine-containing dipeptides with the ferritin nanopore. Five dipeptides including Cys-Phe (CF), Cys-Arg (CR), Cys-Lys (CK), Cys-Asp (CD), Cys-Glu (CE) are detected and we find that CF, CR and CK display homogeneous blockage signals while CD and CE cause no signals (Fig. 5a–d and Supplementary Fig. 20). Negatively charged molecules, as reported by Yang et al., are hard to enter the cavity of ferritin via negatively charged C3 channel due to electrostatic repulsion71, which can explain why the negatively charged CD and CE cause no signals (Supplementary Fig. 21). The signal statistics reveal that all CF, CR, and CK have larger volumes but smaller blockage fractions than L-Cys (Fig. 5e, f). As reported, all the charge, polarity and size of analytes can influence the current blockage fraction in nanopore analysis6,9,10. In our experiments, the blockage fractions of CK, CR, and CF are all smaller than that of L-Cys, with CF having the smallest ΔI/I0, and the difference between CK and CR is minimal (Fig. 5e, f). It is understood that the smaller L-Cys is able to enter into the deeper site of the channel causing larger blockage fraction, while the dipeptides have too large volumes to enter deeper, causing smaller blockage fractions. The CF has the strongest hydrophobicity and largest volume in all of them displaying the smallest blockage fraction. Given that lysine (K) and arginine (R) are hydrophilic polar amino acids, CK and CR are more likely to penetrate deeper into the C3 channel compared to CF, resulting in larger blockage fractions than CF. The charge interaction is believed to dominate in such a strongly negatively charged C3 channel, making it difficult to distinguish between the similarly positively charged CK and CR despite their difference in volume. To decrease the charge interaction between CR/CK and C3 channel, we improve ionic strength by using higher concentration KCl buffer (2 M KCl) in nanopore experiments (Supplementary Fig. 22). In this condition, the blockage fraction is dominated by the volume exclusion effect, which leads to a larger ΔI/I0 for larger CR.
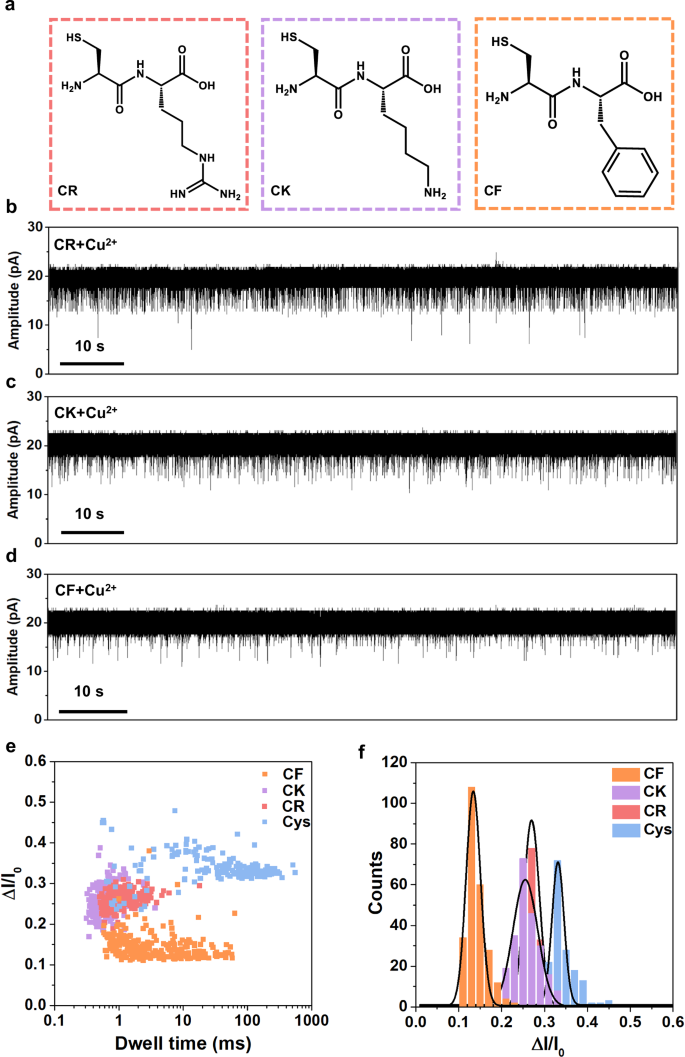
a Structures of CR, CK, and CF dipeptides. b–d Current traces at +40 mV of CR (80 µM), CK (80 µM), and CF (80 µM) in the presence of Cu2+ (80 µM), respectively. e, f Scatter plot and histogram showing the dwell time and ΔI/I0 of blockage signals in dipeptides detection. The signals of cysteine are also displayed for comparison. n = 174, 214, 238, and 251 for Cys, CR, CK, and CF, respectively