Article Content
Introduction
Polycystic ovary syndrome (PCOS) is one of the most common endocrine and metabolic disorders affecting women of reproductive age, with a global prevalence ranging from 5 to 15%. It is clinically characterized by menstrual irregularities, hyperandrogenism, and polycystic ovarian morphology [1, 2]. Beyond reproductive symptoms, PCOS is strongly associated with metabolic disturbances such as insulin resistance, dyslipidemia, obesity, and increased risk of type 2 diabetes and cardiovascular disease [3, 4]. Despite its high prevalence and significant impact on quality of life, its etiology remains unclear, involving a complex interplay of genetic, hormonal, inflammatory, and environmental factors [5, 6].
Recent studies have explored the role of oxidative stress and inflammatory responses in PCOS pathophysiology, suggesting that these processes may impair ovarian function and exacerbate insulin resistance. Notably, emerging literature highlights the relevance of targeted antioxidant therapy in PCOS management [7]. Additionally, recent findings highlight the molecular and epigenetic changes associated with PCOS, emphasizing the need for gene-targeted interventions [8].
Growth differentiation factor 9 (GDF9) and bone morphogenetic protein 15 (BMP15) are oocyte-specific paracrine factors that are essential for folliculogenesis, granulosa cell proliferation, and oocyte maturation [9]. Abnormal expression of these genes contributes to follicular arrest and impaired ovarian development, core features of PCOS [10]. Likewise, phosphatase and tensin homolog (PTEN) is a critical regulator of follicle development and survival, functioning through the PI3K/AKT signaling pathway, and its dysregulation is associated with ovarian dysfunction and cyst formation [11].
Oxidative stress, resulting from an imbalance between reactive oxygen species (ROS) and antioxidant defense mechanisms, plays a critical role in the pathogenesis of PCOS. Elevated ROS levels impair oocyte quality, promote follicular atresia, and compromise ovarian function [12]. Among the antioxidant defense systems, superoxide dismutase (SOD) and glutathione peroxidase (GPx) serve as key enzymes that protect against oxidative damage by neutralizing free radicals and peroxides [13, 14].
Conventional pharmacological interventions, such as clomiphene citrate (Clom) and metformin, are widely used to restore ovulation and improve insulin sensitivity in PCOS patients [15,16,17]. Clomiphene citrate is a selective estrogen receptor modulator that stimulates the hypothalamic-pituitary-ovarian axis and induces ovulation. However, these treatments often have limited efficacy and side effects, which has prompted interest in natural alternatives.
Stevia (Stevia rebaudiana Bertoni), a plant known for its natural sweetening properties, has attracted attention for its antioxidant, anti-inflammatory, and metabolic-modulating effects [18]. Its aqueous extract has demonstrated the ability to reduce oxidative stress, regulate insulin levels, and improve reproductive parameters in experimental models [19].
Therefore, this study aimed to evaluate the effects of aqueous Stevia rebaudiana extract (SAE) and clomiphene citrate on the expression of GDF9, PTEN, and BMP15 genes, as well as on antioxidant status, in a letrozole-induced PCOS model in Wistar rats. This investigation provides insight into the molecular and oxidative pathways involved in PCOS and explores the therapeutic potential of combining natural and pharmaceutical interventions.
Methods
Extraction of aqueous extract from Stevia rebaudiana leaves
Dried leaves of Stevia rebaudiana Bertoni were extracted using hot water at 65 °C, with a leaf-to-water ratio of 1:45 (w/v). The extraction lasted for three hours, after which the crude extract was filtered through Whatman No. 4 filter paper. To purify the extract, 5% calcium hydroxide [Ca(OH)₂] was added based on the dry leaf weight (repeated twice). The resulting filtrates were passed through an Amberlite IR-4B resin ion-exchange column (Sigma-Aldrich, USA) to remove pigmentation. After 24 h at room temperature, the filtrate was filtered and concentrated using a rotary vacuum evaporator (Jeio Tech, Model Cubic SDC-30/30U, South Korea) [20, 21].
Animals
Fifty adult female Wistar rats (180–220 g) were purchased from the Animal House, Faculty of Medicine, Rafsanjan University of Medical Sciences (RUMS). Animals were maintained under a 12:12 h light/dark cycle at 20–25 °C with free access to food and water. Animal procedures followed the NIH Guide for the Care and Use of Laboratory Animals.
Estrous cycle determination and induction of PCOS
To synchronize the estrous cycle, rats received 100 µg of estradiol valerate (Aburaihan Co., Iran) in 0.2 ml of olive oil intramuscularly. After 42 h, 50 µg of progesterone (Aburaihan Co., Iran) was injected intramuscularly. Vaginal smears were collected 6 h later and stained with crystal violet (Merck, Germany) for microscopic analysis (100×).
Letrozole (1 mg/kg/day; Iran Hormone, Iran) was administered orally for 21 days, dissolved in 0.5% carboxymethyl cellulose (CMC; Merck, Germany) to induce PCOS [8, 22]. PCOS was confirmed by vaginal smear analysis showing prolonged diestrus and cycle irregularity.
Grouping of animals
Rats were randomly divided into 5 groups (n = 10 per group):
Group 1 (Control): 1 ml of CMC orally for 49 days.
Group 2 (PCOS): Letrozole (1 mg/kg) orally for 21 days, followed by CMC for 28 days.
Group 3 (SAE): Letrozole for 21 days, then SAE (400 mg/kg) orally for 28 days.
Group 4 (Clom): Letrozole for 21 days, then clomiphene citrate (Clom, 100 mg/kg) (Razak Labs, Iran) for 28 days.
Group 5 (CS): Letrozole for 21 days, followed by a combination of Clom (100 mg/kg) and SAE (400 mg/kg) for 28 days.
Measurement of antioxidant and hormonal parameters
At study end, rats were anesthetized with ketamine (80 mg/kg) and xylazine (10 mg/kg) (Alfasan, Netherlands). Blood was collected from the retro-orbital sinus. Serum was separated and stored at − 80 °C.
Hormone levels (LH, FSH, estrogen, progesterone) were measured using Pishtaz Teb ELISA kits (Pishtaz Teb Diagnostics, Iran; Kit Nos: LH-E2010, FSH-E1010, E2-E2210, PRG-E2310).
SOD and GPx activities were measured using ZellBio kits (Germany; Kit Nos: ZB-SOD96A, ZB-GPX96A).
Histopathology
Both ovaries were excised, fixed in 10% buffered formalin, and stored at 4 °C. One ovary per animal was processed, embedded in paraffin, sectioned at 5 μm, and stained with hematoxylin and eosin (H&E) for microscopic evaluation. Histological structures including antral follicles, corpus luteum, atretic and cystic follicles were identified using light microscopy. Quantification of follicles was conducted in 3 randomly selected sections per ovary, and counting was done per entire section under 100× magnification. The number of follicles, including primordial follicles, was estimated using the physical dissector stereological method based on the entire ovary. In this approach, consecutive tissue sections were prepared and stained, and follicles were counted using a Nikon microscope. For each ovary, systematic random sampling was applied. Two consecutive sections were projected simultaneously -one as the reference and the other as the lookup section- and 13 × 13 mm counting frames were randomly placed on the images. Only oocytes that did not touch the exclusion line and were absent in the lookup section were counted. The total number of follicles in the whole ovary was then calculated using stereological formulas, accounting for the reference volume obtained by the Cavalieri principle. We have revised the Materials and Methods section to reflect this clarification and improve the reproducibility of the methodology [23].
The number of follicles was calculated using the following formula: N = Nv × V(Ref).
where N is the total number of follicles, Nv is the numerical density of follicles per unit volume, and V(Ref) is the reference volume of the tissue, which was determined using the Cavalieri principle: V = ∑P × a(p)Xt.
In this formula, V represents the volume, ∑P is the total number of points hitting the region of interest, a(p) is the area associated with each point, and t is the thickness of the sections. The term a(p) refers to the volume of space surrounding a single intersection point.
RNA extraction and cDNA synthesis
Ovaries preserved in liquid nitrogen were powdered and RNA was extracted using Parstous RNA Extraction Kit (Parstous Biotech, Iran). RNA quality was confirmed with a Nanodrop (Thermo Fisher Scientific, USA) and 1% agarose gel electrophoresis.
1 µg of RNA was reverse-transcribed using Parstous cDNA Synthesis Kit (Parstous Biotech, Iran).
RT-PCR was conducted using SYBR Green Master Mix (Amplicon, Denmark) with the following primers:
Gdf9 (F: AGCCACTTACAGCATCCTTC / R: TCCAGTTGTCCCATTTGAGC).
Bmp15 (F: GAGGTCCTTGGCATATACAG / R: GATGAAGTTGATGGCGGTAG).
Pten (F: ATACCAGGACCAGAGGAAACC / R: TTGTCATTATCCGCACGCTC).
Actb (F: CGAGTACAACCTTCTTGCAGC / R: TCAGGGTCAGGATGCCTCTC).
Relative gene expression was calculated using the 2^(-ΔΔCt) method.
Statistical analysis
Data were analyzed using SPSS version 21. One-way ANOVA followed by Tukey’s post hoc test was used for multiple comparisons. Data are presented as mean ± SD, with P < 0.05 considered statistically significant. Normality of data distribution was assessed using the Shapiro-Wilk test.
Ethical consideration
All experimental procedures were approved by the Ethics Committee of Rafsanjan University of Medical Sciences (ID: IR.RUMS.REC.1401.235) and followed national animal care guidelines.
Results
Letrozole-induced PCOS and its effects on the estrous cycle
The induction of PCOS using letrozole (1 mg/kg for 21 days) was confirmed by significant alterations in the estrous cycle of female Wistar rats. Before induction, all rats exhibited regular estrous cycles, characterized by defined and alternating phases of proestrus, estrus, metestrus, and diestrus. However, letrozole administration disrupted this pattern, leading to prolonged diestrus phases, irregular cycles, and anovulation (Fig. 1A–D).
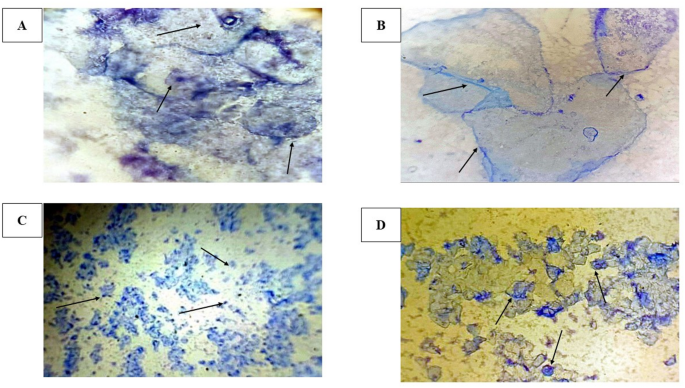
Representative cytological images of crystal violet-stained vaginal smears illustrating the estrous cycle phases in female Wistar rats at 100x magnification. (A) Proestrus, characterized by a predominance of nucleated epithelial cells; (B) Estrus, exhibiting a predominance of cornified epithelial cells; (C) Diestrus, marked by a predominance of leukocytes; (D) Metestrus, displaying a mixture of leukocytes and epithelial cells
Body weight changes in laboratory rats during the treatment period
On day 1, there were no significant differences in baseline body weights among the groups (p = 0.82). By day 22 (post-induction), the PCOS, SAE, Clom, and CS groups exhibited significantly higher body weights compared to the control group (p < 0.01). On day 49 (post-treatment), the PCOS group maintained the highest weight (251.40 ± 6.69 g, p < 0.01 vs. control), whereas the SAE (242.30 ± 7.13 g), Clom (238.30 ± 4.90 g), and CS (233.70 ± 3.20 g) groups showed a significant decrease compared to the PCOS group (p < 0.01, Table 1).
Effect of treatments on serum hormone levels
Figure 2A–E shows hormone level comparisons. The PCOS group had a significant increase in LH (p < 0.0001) and LH/FSH ratio (p < 0.0001), and a decrease in FSH (p < 0.0001) compared to the control group. Treatment with SAE, Clom, and CS significantly reduced LH and LH/FSH ratio while increasing FSH compared to PCOS (p < 0.0001 for all comparisons).
Estrogen levels were significantly lower in the PCOS group (p < 0.0001) but improved in all treatment groups (p < 0.0001 vs. PCOS).
Progesterone was reduced in all PCOS-induced groups compared to control (p < 0.0001), but SAE, Clom, and CS significantly elevated progesterone versus PCOS (p < 0.0001).
Clom and CS had higher progesterone levels than SAE (p < 0.0001), while CS was slightly lower than Clom (p < 0.0001).
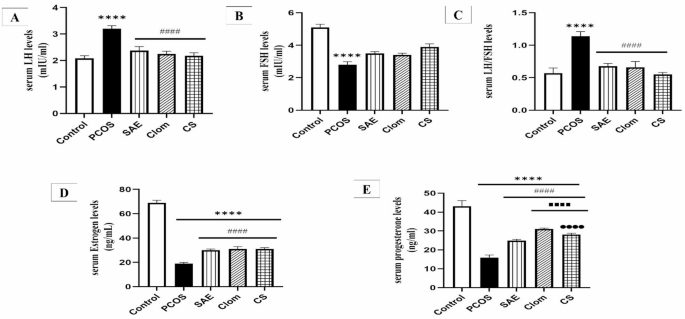
Comparison of serum LH (A), FSH (B), LH/FSH (C), estrogen (D), and progesterone (E) levels in PCOS rats treated with SAE, Clom, and CS. Values are presented as mean ± SD, with n = 10 rats per group. Statistical analysis was performed using one-way analysis of variance (ANOVA) followed by Tukey’s post-hoc test. ****P < 0.0001, significantly different from the control group; ####P < 0.0001, significantly different from the PCOS group; ▪▪▪▪P < 0.0001, significantly different from the SAE group; ●●●●P < 0.0001, significantly different from the Clom group. PCOS, polycystic ovary syndrome; SAE, stevia aqueous extract; Clom, clomiphene; CS, SAE/Clom combination
Gene expression alterations of PTEN, GDF9, and BMP15
Gene expression of BMP15, GDF9, and PTEN is presented in Fig. 3. BMP15 and GDF9 expression were significantly higher in the PCOS group compared to the control (p < 0.001). These expressions were significantly reduced in SAE, Clom, and CS groups versus PCOS (p < 0.001). PTEN was significantly reduced in PCOS and SAE groups compared to control (p < 0.001). Clom and CS significantly increased PTEN expression vs. SAE (p < 0.001).
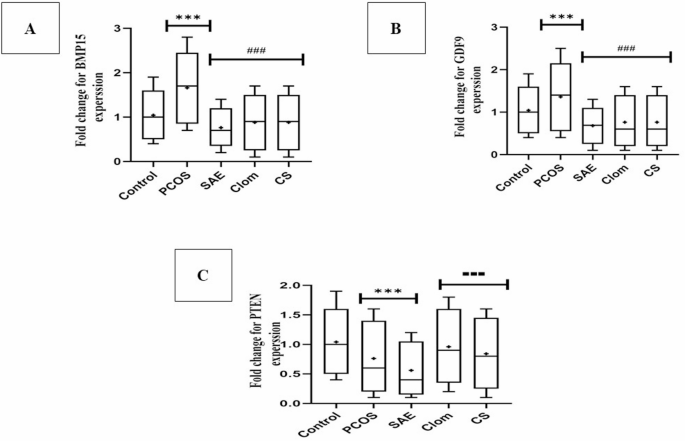
Effects of PCOS and treatment interventions on the expression of BMP15 (A), GDF9 (B), and PTEN (C) genes in ovarian tissue. Data are presented as mean ± SD (n = 10 per group). Statistical analysis was performed using one-way ANOVA followed by Tukey’s post hoc test. ***P < 0.001 vs. Control group; ###P < 0.001 vs. PCOS group; ▪▪▪P < 0.001 vs. SAE group. PCOS, Polycystic Ovary Syndrome; SAE, Stevia Aqueous Extract; Clom, Clomiphene Citrate; CS, SAE/Clom Combination
Comparative analysis of antioxidant enzyme levels (SOD and GPX)
As shown in Table 2, the PCOS and SAE groups had significantly lower SOD levels than the control (p < 0.001). Clom and CS treatments significantly increased SOD compared to PCOS (p < 0.001). GPX levels were also reduced in the PCOS group (p < 0.001) but were significantly elevated in the SAE, Clom, and CS groups compared to PCOS (p < 0.001).
Histological features of letrozole-induced PCOS
H&E-stained ovarian sections from PCOS rats (Fig. 4A–C) showed multiple cystic follicles, reduced corpora lutea, and hyperplasia of the theca layer.
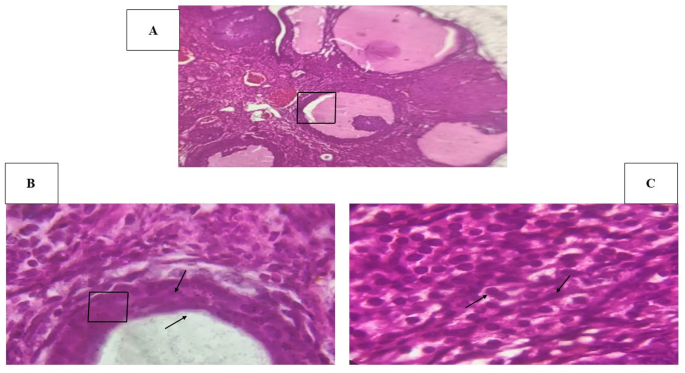
Histological features of ovarian tissue in a PCOS model (H&E staining, 100× magnification). (A) Ovarian section showing multiple large cystic follicles and reduced presence of corpora lutea, indicating anovulation and follicular arrest. (B) Higher magnification of a cystic follicle wall, demonstrating the presence of a thickened theca layer and vascular changes, potentially reflecting luteinization and stromal hyperplasia. C) Detailed view of stromal tissue with evident theca cell hyperplasia, characterized by increased cell density and nuclear alterations, suggestive of endocrine disruption commonly associated with PCOS pathology
Histological observations in treatment groups
Histological observations across the treatment groups are depicted in Fig. 5A–E. Control ovaries displayed normal follicles, antral follicles, and corpora lutea. In contrast, PCOS ovaries exhibited dominant cystic structures with a complete absence of ovulation markers. Ovaries from SAE-treated rats revealed a combination of atretic follicles, normal vessels, and corpora lutea. Clom-treated ovaries showed partial restoration, with emerging follicles and corpora lutea. Finally, CS-treated ovaries demonstrated improved follicular integrity, although some persistent atresia was observed, indicating partial synergy between the SAE and Clom treatments.
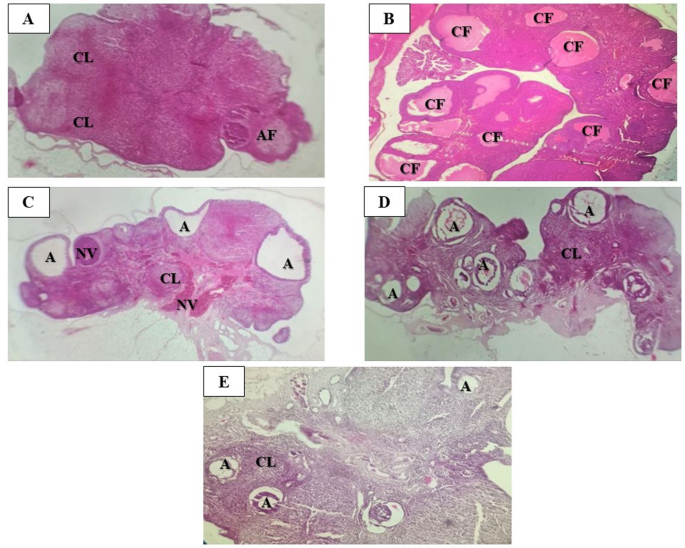
Histological changes in ovarian tissue stained with Hematoxylin and Eosin (H&E) at 100x magnification. (A) Control; (B) PCOS; (C) SAE; (D) Clom; (E) CS. Key observations include corpora lutea (CL), normal vessels (NV), antral follicles (AF), atretic cystic follicles (A), and cystic follicles (CF). PCOS, polycystic ovary syndrome; SAE, stevia aqueous extract; Clom, clomiphene; CS, SAE/Clom combination
Quantitative analysis of ovarian follicles
Table 3 summarizes follicular counts. PCOS rats had significantly fewer follicles across all categories (except cystic follicles), compared to control (p < 0.001). SAE, Clom, and CS treatments significantly increased the number of corpus luteum and reduced cystic follicles compared to PCOS (p < 0.001).
Discussion
This study provides compelling evidence for the efficacy of aqueous Stevia rebaudiana extract (SAE), clomiphene citrate (Clom), and their combination (CS) in ameliorating the multifaceted pathophysiology of polycystic ovary syndrome (PCOS) in a letrozole-induced Wistar rat model. The experimental outcomes reaffirm PCOS as a condition not solely limited to reproductive dysfunction but also involving significant metabolic, hormonal, genetic, and oxidative disruptions. Notably, the combined therapeutic strategy (CS) demonstrated additive or synergistic benefits across most parameters evaluated, supporting an integrative treatment model that leverages both pharmaceutical and phytotherapeutic agents. This combination may be particularly relevant in clinical scenarios where monotherapy fails or where adjunct support is necessary to address the metabolic or oxidative dimensions of PCOS.
The letrozole-induced model successfully replicated key pathological traits of human PCOS, including hormonal imbalance, disrupted estrous cycles, and cystic follicle formation, consistent with previous models [24, 25]. By inhibiting aromatase, letrozole disrupts estrogen biosynthesis, leading to elevated androgen levels. This hormonal environment disturbs hypothalamic feedback, impairs follicular development, and causes ovulatory dysfunction. The observed prolonged diestrus and irregular estrous cycles in this study confirm the functional disruption of the hypothalamic-pituitary-ovarian (HPO) axis [26]. This makes the model suitable for investigating not only therapeutic outcomes but also underlying pathophysiological mechanisms of PCOS.
Body weight gain in the PCOS group, along with elevated LH and reduced FSH, reflects metabolic and reproductive dysfunction. Treatments with SAE, Clom, and CS effectively reduced weight and restored hormonal balance by day 49, possibly via improvements in insulin sensitivity, estrogen regulation, and androgen suppression, which aligns with prior findings [27,28,29]. Although weight reduction was not the primary aim, it may indirectly reflect improvements in insulin sensitivity or lipid metabolism, especially since Stevia rebaudiana is known to modulate glucose and lipid profiles. Letrozole-induced hyperandrogenism can increase visceral fat deposition and insulin resistance, contributing to weight gain. The greater weight reduction in CS-treated rats suggests a potential metabolic synergy that warrants further exploration with glucose tolerance and insulin resistance assessments. In this context, SAE may act as an insulin-sensitizing agent through bioactive compounds like stevioside, which enhance glucose uptake and modulate hepatic gluconeogenesis [30].
Progesterone levels, suppressed in PCOS due to defective luteal activity, were significantly improved post-treatment. Clom and CS showed superior restoration compared to SAE, reflecting the well-established ovulation-inducing effects of Clom, while SAE’s impact may stem from phytoestrogens and glycosides modulating hypothalamic feedback [31]. Progesterone restoration serves as an indirect biomarker for successful ovulation and corpus luteum formation. Given that progesterone plays a crucial role in endometrial receptivity and early pregnancy maintenance, the observed improvements suggest a positive trajectory for potential fertility outcomes. Interestingly, while CS improved progesterone, some histological evidence of incomplete luteinization suggests the need for dose optimization or adjunctive luteal phase support. It is possible that although ovulation was triggered, luteal phase hormonal support might still be insufficient in some cases. This is consistent with literature suggesting that clomiphene-induced ovulation does not always equate to a fully functional luteal phase [32].
Elevated GDF9 and BMP15 expression in PCOS confirms their role in follicular arrest and impaired oocyte maturation [33, 34]. Treatments reduced their expression, supporting follicular normalization. Interestingly, PTEN, downregulated in PCOS, was notably restored by Clom and CS, suggesting a mechanism involving the PI3K/AKT pathway and granulosa cell survival [35]. GDF9 and BMP15, although essential for normal folliculogenesis, appear to be dysregulated in PCOS, potentially due to aberrant upstream endocrine cues. Their overexpression may disrupt the delicate granulosa-oocyte signaling necessary for ovulation, leading to cystic degeneration. PTEN, a negative regulator of the PI3K/AKT pathway, is vital in maintaining a balance between follicular dormancy and activation. Its reduced expression in PCOS ovaries may result in premature or aberrant follicle growth. Restoration of PTEN by Clom and CS may reflect improved granulosa cell survival, regulated follicular recruitment, and reduced follicular atresia. Moreover, PTEN is increasingly being recognized for its role in regulating oocyte quality, mitochondrial dynamics, and ovarian reserve -dimensions critically affected in PCOS [36].
Oxidative stress plays a major role in PCOS. Our results show significantly reduced SOD and GPX in PCOS ovaries. SAE, Clom, and CS treatments restored these levels, with CS being most effective. SAE’s effect is attributed to flavonoids, phenolic acids, and stevioside derivatives, which scavenge ROS and improve mitochondrial efficiency [18, 19, 37]. This supports the hypothesis that oxidative stress not only exacerbates PCOS symptoms but may actively participate in its etiology by impairing oocyte quality, disrupting steroidogenesis, and inducing inflammatory cascades. The antioxidant effect of SAE likely extends beyond direct free radical neutralization, possibly influencing the expression of redox-sensitive genes such as Nuclear factor erythroid 2-related factor 2 (NRF2) and Sirtuin 1 (SIRT1). Clom and CS treatments also significantly boosted antioxidant enzyme levels, suggesting an indirect enhancement of cellular antioxidant capacity -possibly via improved endocrine balance, reduced inflammation, or modulation of mitochondrial biogenesis. These improvements may not only restore ovarian health but also enhance oocyte quality and embryonic competence, critical for successful reproduction [38].
Histologically, PCOS ovaries displayed theca cell hyperplasia, cystic follicles, and absent corpora lutea, while treated groups showed partial to near-complete restoration of follicular morphology, especially with CS. These findings align with studies on hormonal and metabolic corrections in PCOS rats treated with phytocompounds and ovulation agents [39]. The observed increase in antral follicles and corpora lutea in CS-treated rats strongly suggests re-established ovulation and follicular maturity. Restoration of corpus luteum formation is particularly significant, as it marks the end-point of successful follicular development and confirms the transition from a dysfunctional to a functional ovary. Nevertheless, residual atresia and incomplete luteinization in some specimens highlight inter-individual variability or perhaps limitations in treatment duration or dosing strategy. These subtle histological inconsistencies could also reflect asynchronous follicular development or the presence of unresolved systemic metabolic stress [40].
The dual modulation of hormonal and oxidative parameters by SAE and Clom offers a multifaceted approach to PCOS management. While Clom remains a gold standard for ovulation induction, its limited impact on metabolic and oxidative features is well documented. SAE, by contrast, complements this gap through its antioxidative and potentially insulin-sensitizing properties. The combination strategy may, therefore, represent a novel paradigm for treating both reproductive and metabolic dimensions of PCOS, reducing reliance on polypharmacy. This integrative strategy may be especially relevant in women with clomiphene resistance, metabolic syndrome, or contraindications to conventional pharmacotherapy [41].
Further studies are needed to establish dose–response relationships, explore underlying pathways (e.g., insulin signaling, AMPK activation), investigate long-term fertility outcomes, and evaluate the anti-inflammatory effects and immune modulation by SAE. Inclusion of parameters such as serum insulin, HOMA-IR index, inflammatory cytokines (e.g., IL-6, TNF-α), and mitochondrial markers could provide a more comprehensive picture. Additionally, omics-based approaches like transcriptomics or metabolomics could offer deeper insight into the global impact of these interventions at a systems biology level. Clinical translation would benefit from safety profiling, pharmacokinetic studies, and controlled human trials to verify efficacy, particularly in populations with varied phenotypic expressions of PCOS.
Limitations
This study used only a single dose per treatment, limiting assessment of dose–response relationships. Key metabolic and inflammatory markers, such as insulin resistance and cytokine levels, were not evaluated. Fertility outcomes and long-term effects post-treatment remain unknown. Additionally, while the letrozole-induced rat model reflects human PCOS features, species differences limit direct clinical translation. Future studies should explore varying doses, assess metabolic and reproductive endpoints, and consider translational research in human subjects.
Conclusion
This study established a reliable letrozole-induced PCOS model in Wistar rats, effectively replicating key features of human PCOS, including hormonal imbalances, ovarian dysfunction, and metabolic disturbances. Treatment with SAE, Clom, and their combination (CS) significantly improved hormonal balance, antioxidant defenses, and ovarian histology. Clom and CS exhibited superior efficacy in promoting follicular development and ovulation, while also reducing oxidative stress. SAE provided moderate benefits, whereas CS demonstrated synergistic effects in addressing both hormonal and metabolic challenges. These findings offer valuable insights into potential therapeutic strategies for managing PCOS.